
Guimaraesiella ( Dicrurobates ) Gustafsson & Bush, 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4885.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:081203D8-39FF-41C3-A79A-BB63F47AB3B1 |
|
DOI |
https://doi.org/10.5281/zenodo.4332162 |
|
persistent identifier |
https://treatment.plazi.org/id/3487B0CA-0E09-4473-AB17-8DBB7D75DEA2 |
|
taxon LSID |
lsid:zoobank.org:act:3487B0CA-0E09-4473-AB17-8DBB7D75DEA2 |
|
treatment provided by |
Plazi |
|
scientific name |
Guimaraesiella ( Dicrurobates ) Gustafsson & Bush |
| status |
subgen. nov. |
Guimaraesiella ( Dicrurobates) Gustafsson & Bush , new subgenus
urn:lsid:zoobank.org:act:
Brueelia Kéler, 1936: 257 ( in part).
Brueelia “clade C” Bush et al. 2016: 743, fig. 3c.
Type species: Brueelia dicruri Ansari, 1955 ex Dicrurus macrocercus albirictus (Hodgson, 1836) View in CoL .
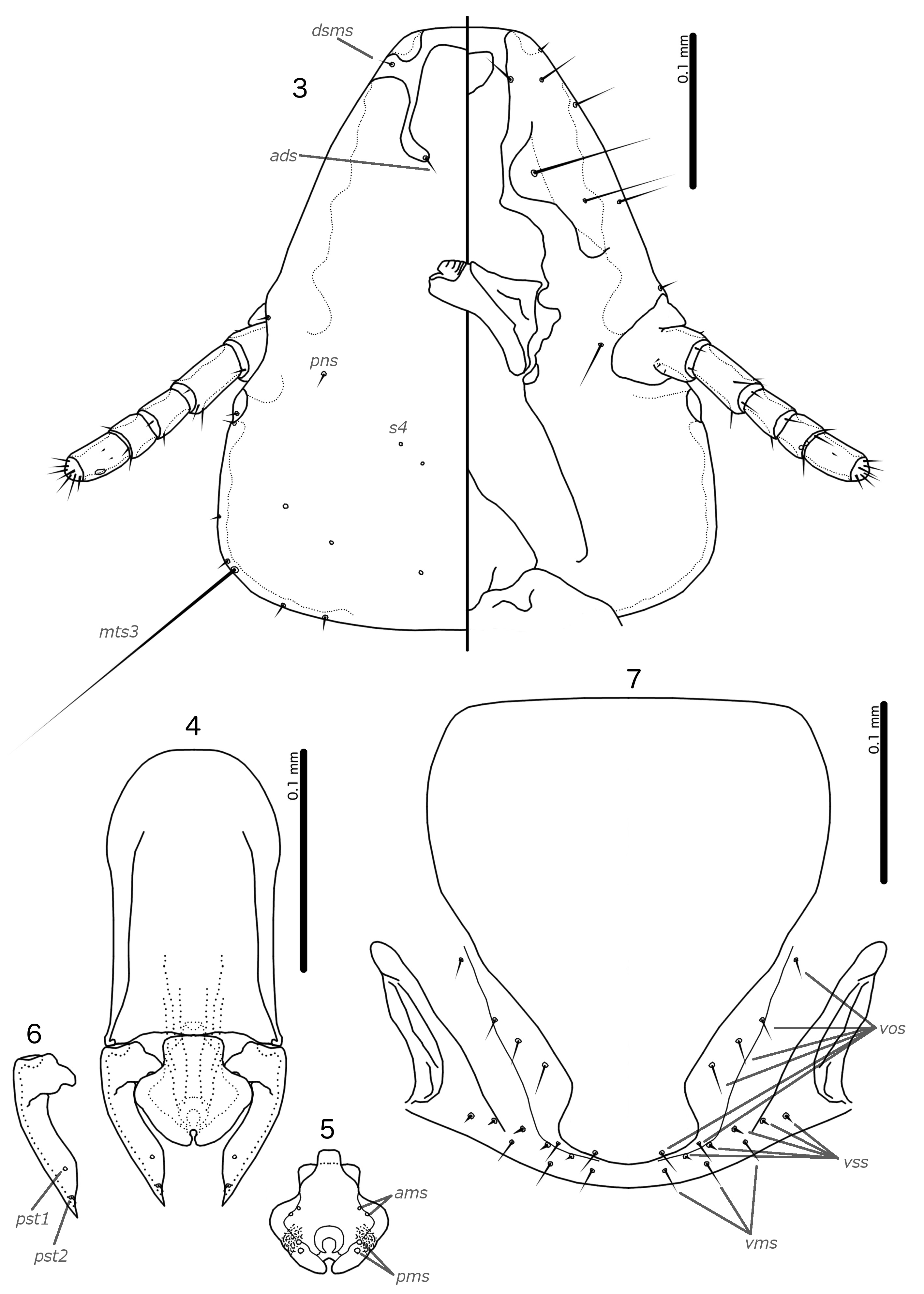
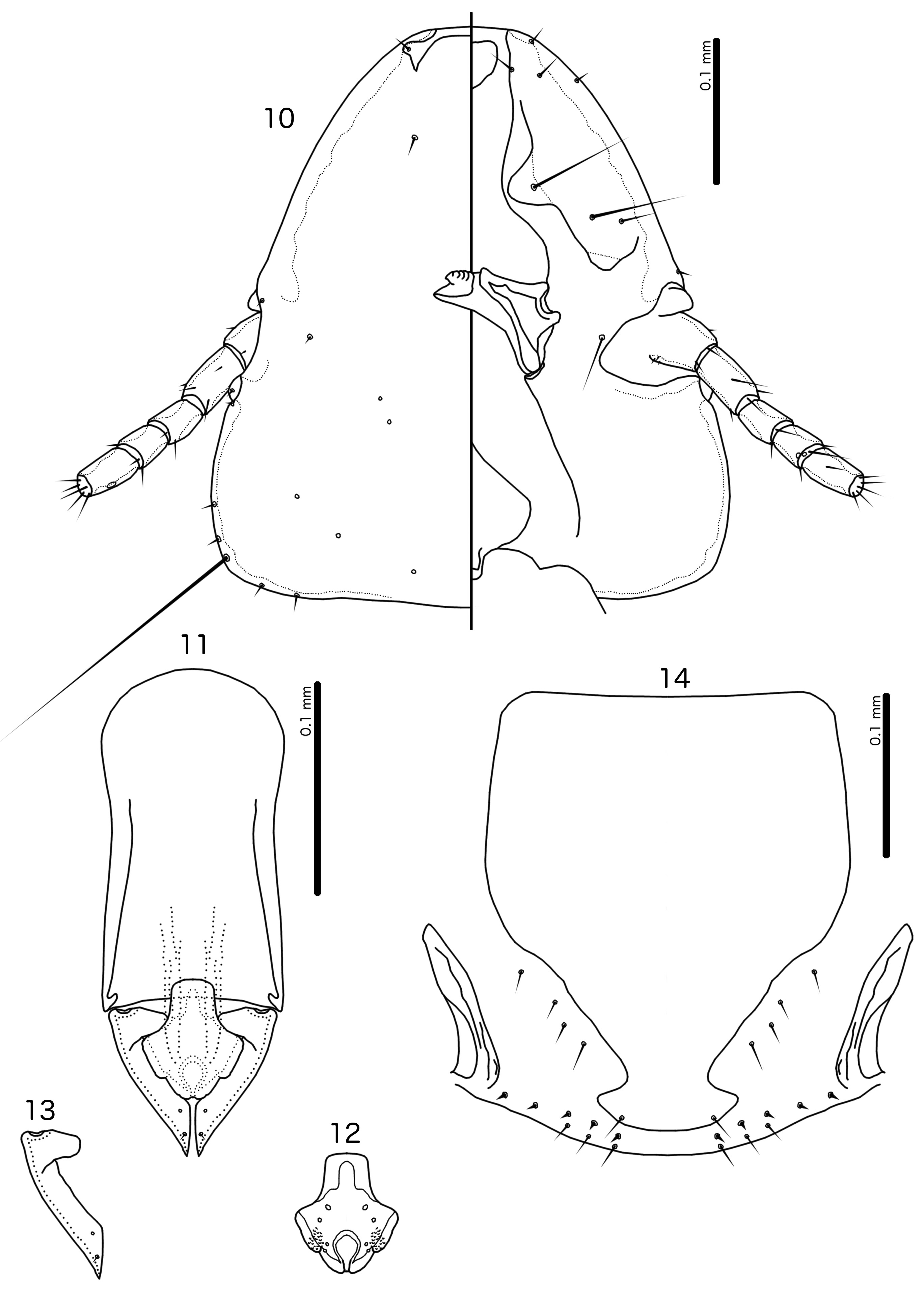
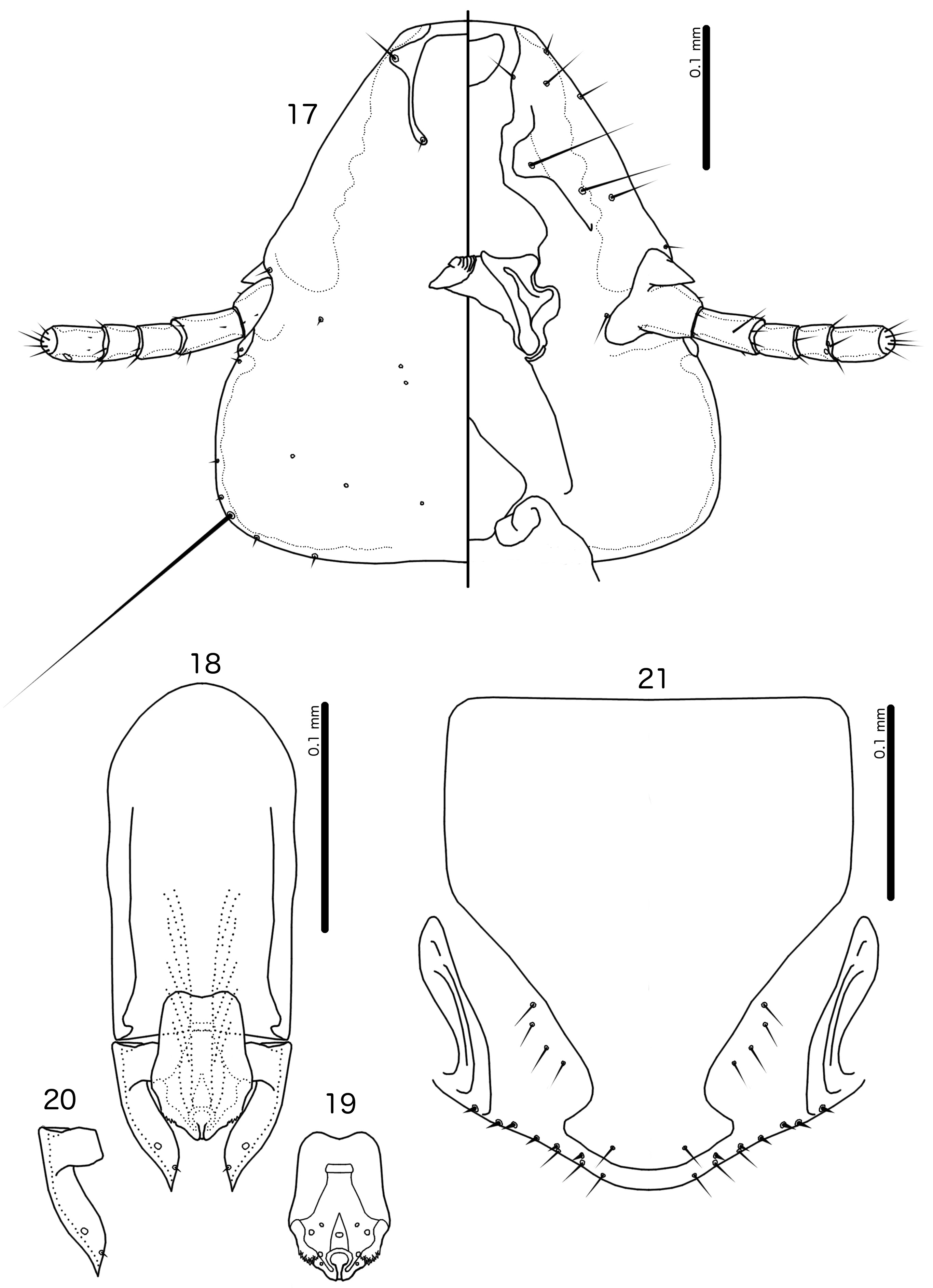
Diagnosis. The subgenus Guimaraesiella ( Dicrurobates) shares the following characters with the nominate subgenus: dorsal preantennal suture present; marginal carina interrupted at least medianly; pns and s4 present; as3 absent; psps present on tergopleurites IV–VII; setal rows absent on all tergopleurites in both sexes; ss present on tergopleurites II–VIII in both sexes; aps present on male tergopleurite VII; parameral heads folded medianly; gonopore open distally. However, species included in Guimaraesiella ( Dicrurobates) can be separated from those in the nominate subgenus by two characters of the male genitalia: (1) gonopore positioned terminally in species of Guimaraesiella ( Guimaraesiella) , but subterminally in species of Guimaraesiella ( Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ); (2) rugose nodi present in species of Guimaraesiella ( Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ), but absent in species of Guimaraesiella ( Guimaraesiella) . Both these characters are found in at least some species included in the subgenera Guimaraesiella ( Mohoaticus) Mey, 2017 and Guimaraesiella ( Cicchinella) Gustafsson et al., 2019a ( gombakensis and tenella species groups). Hence, Guimaraesiella ( Dicrurobates) may be closer to one or both of these subgenera than to Guimaraesiella sensu stricto. Notably, in the phylogeny of Bush et al. (2016: 742, fig. 3(b), clade A-5), the subgenus Guimaraesiella ( Cicchinella) was not placed close to the nominate subgenus; however, no member of the subgenus Guimaraesiella ( Mohoaticus) was represented in that phylogeny, and relationships among the deeper nodes within Guimaraesiella sensu lato were poorly resolved.
Species of Guimaraesiella ( Dicrurobates) can be separated from species of Guimaraesiella ( Cicchinella) with rugose nodi by the following characters: (1) female subgenital plate with cross-piece in Guimaraesiella ( Cicchinella) ( gombakensis and tenella species groups), but without cross-piece in Guimaraesiella ( Dicrurobates) (e.g. Figs 7 View FIGURES 3–7 , 14 View FIGURES 10–14 , 21 View FIGURES 17–21 ); (2) male gonopore ventral, near center of mesosome in Guimaraesiella ( Cicchinella) (both species groups), but subterminal in Guimaraesiella ( Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ); (3) ventral sclerite absent in Guimaraesiella ( Cicchinella) ( gombakensis species group), but present in Guimaraesiella ( Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ); (4) parameral heads with corrugated section in Guimaraesiella ( Cicchinella) (both species groups), but without such corrugation in Guimaraesiella ( Dicrurobates) (e.g. Figs 6 View FIGURES 3–7 , 13 View FIGURES 10–14 , 20 View FIGURES 17–21 ); (5) dorsal preantennal suture completely separating dorsal anterior plate in Guimaraesiella ( Cicchinella) ( tenella species group), but not separating the dorsal anterior plate in Guimaraesiella ( Dicrurobates) (e.g. Figs 3 View FIGURES 3–7 , 10 View FIGURES 10–14 , 17 View FIGURES 17–21 ).
Species of Guimaraesiella ( Mohoaticus) can be separated from Guimaraesiella ( Dicrurobates) by the following characters: (1) dorsal preantennal suture not medially continuous median to ads and dorsal anterior plate continuous with roof of head in Guimaraesiella ( Dicrurobates) (e.g. Figs 3 View FIGURES 3–7 , 10 View FIGURES 10–14 , 17 View FIGURES 17–21 ), but suture medially continuous median to ads and dorsal anterior plate separated from roof of head in Guimaraesiella ( Mohoaticus) ; (2) mesosomal lobes with straight or bulging lateral margins and rugose nodi either poorly delimited or on bulge in Guimaraesiella ( Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ), but with deeply sinuous lateral margins and rugose nodi clearly delimited ventrally by a noticeable ridge in Guimaraesiella ( Mohoaticus) ; (3) gonopore smooth anteriorly in Guimaraesiella ( Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ), but serrated anteriorly in Guimaraesiella ( Mohoaticus) .
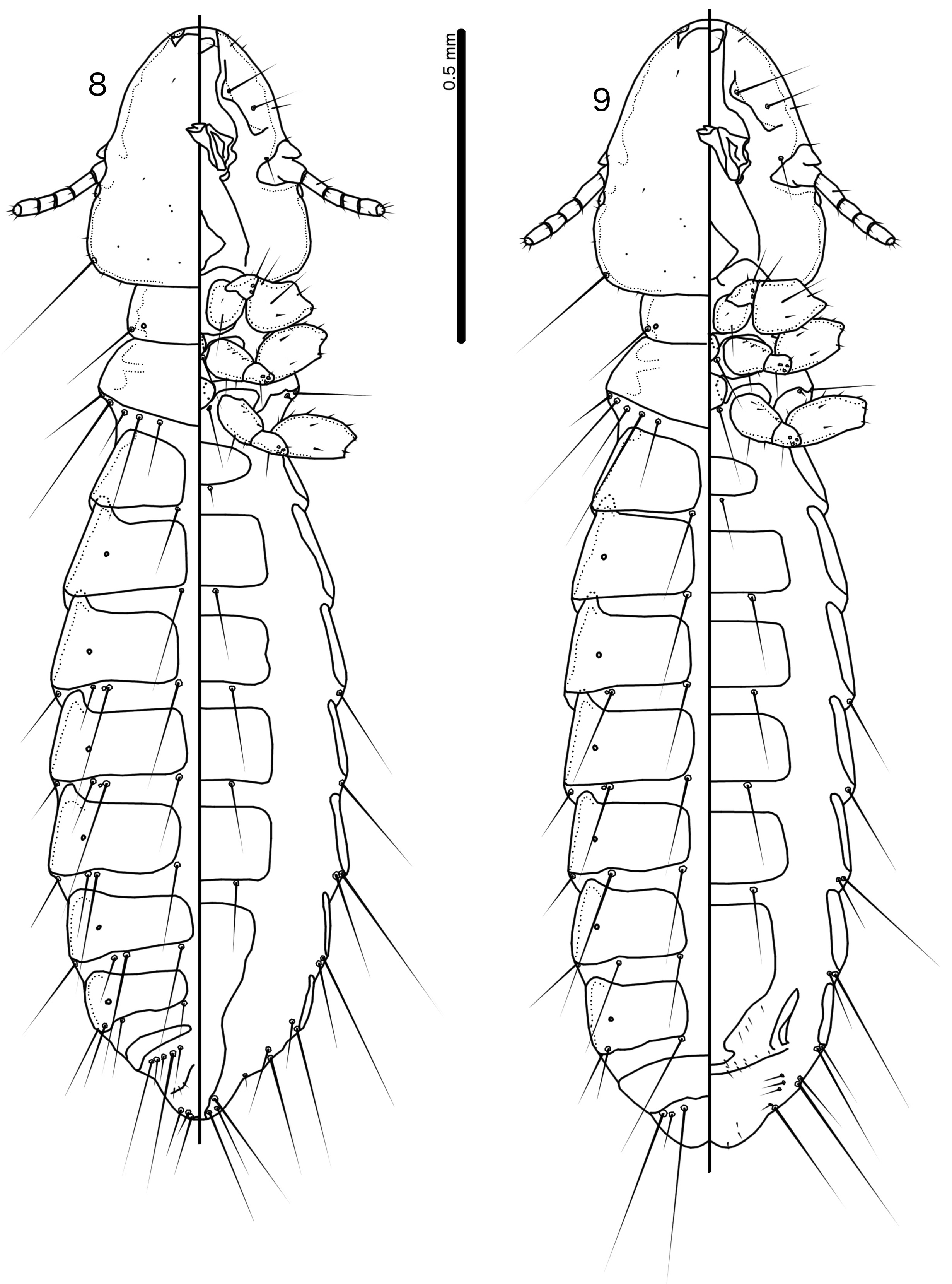
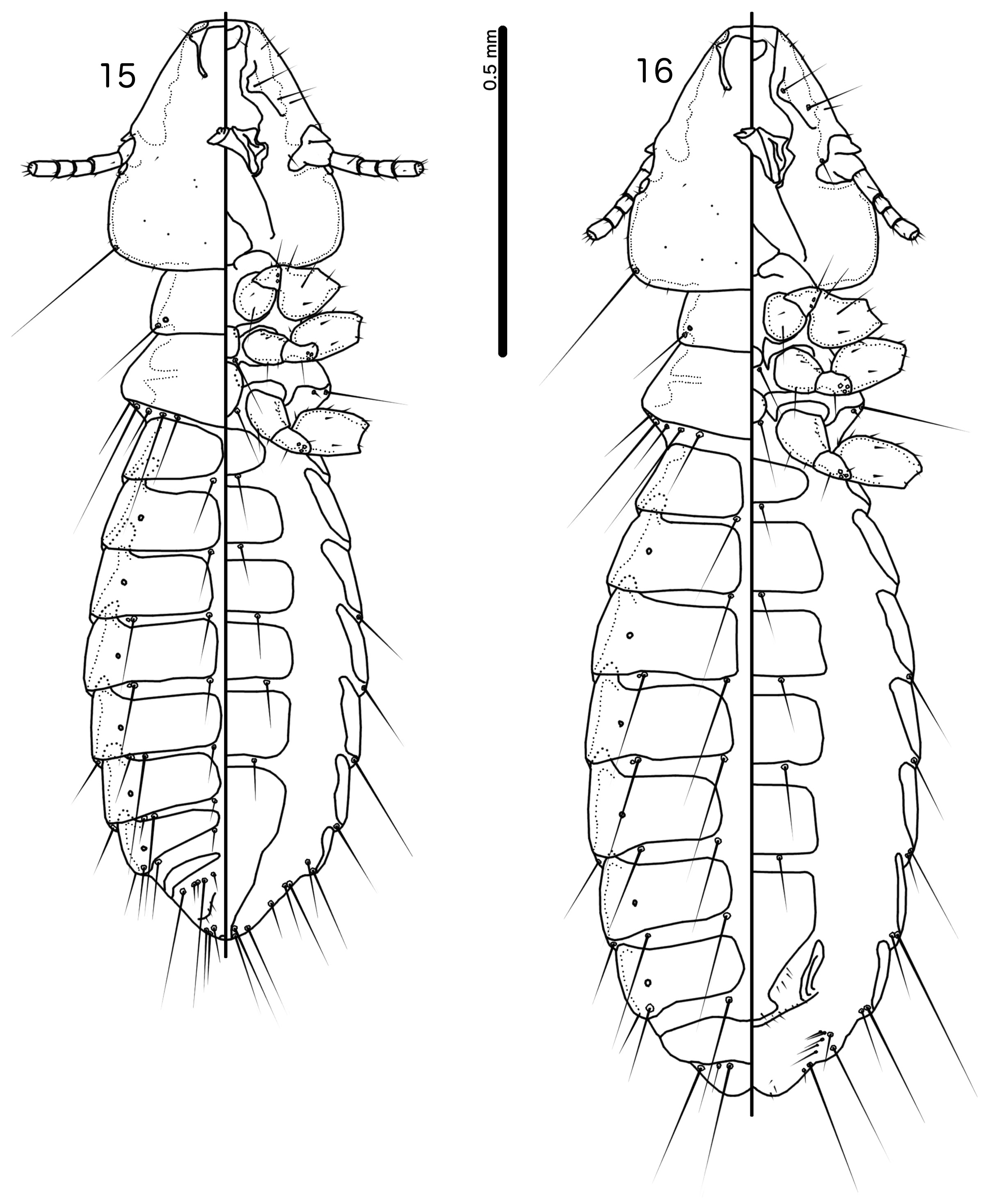
Description. Both sexes. Head shape variable, but preantennal area typically long and roughly trapezoidal, with flattened frons (e.g. Figs 3 View FIGURES 3–7 , 10 View FIGURES 10–14 , 17 View FIGURES 17–21 ). Marginal carina broad, with irregular inner margins, interrupted medianly but not laterally (except in species where dorsal preantennal suture reaches lateral margins of head); frons hyaline, continuous with dorsal preantennal suture which reaches at least dsms, and may reach ads and lateral margins of head; ventral anterior plate present; temporal and occipital carinae not visible; antennae sexually monomorphic; temples gently rounded. Head chaetotaxy as in e.g. Figs 3 View FIGURES 3–7 , 10 View FIGURES 10–14 , 17 View FIGURES 17–21 ; as3 absent; mts3 only temporal macrosetae.
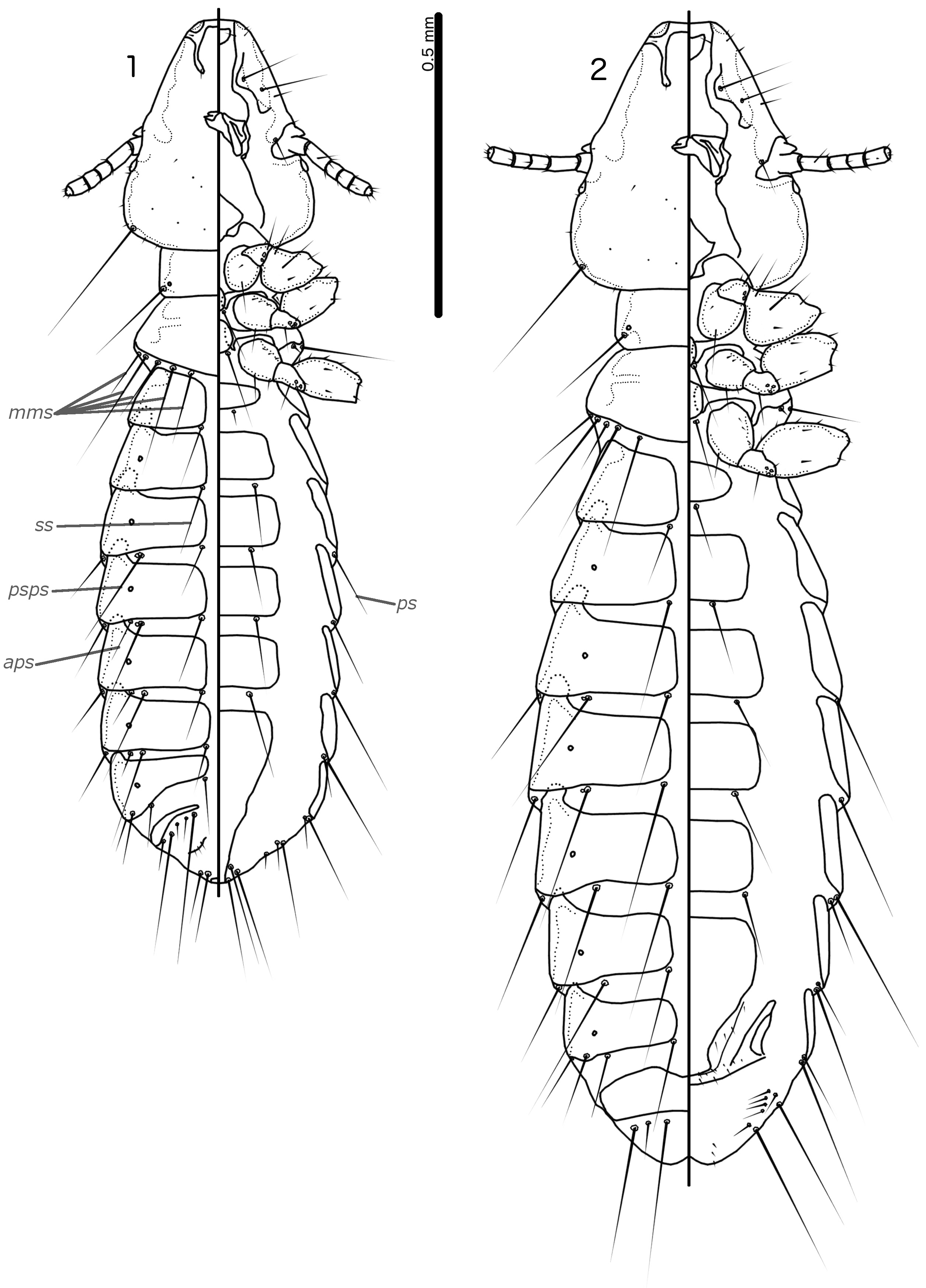
Prothorax rectangular; psps on postero-lateral corners. Pterothorax roughly pentagonal, with lateral margins divergent and posterior margin either rounded or convergent to median point; mms moderately separated medianly. Meso- and metasterna not fused, each with 1 seta on each side on postero-lateral corners (e.g. Figs 1–2 View FIGURES 1–2 , 8–9 View FIGURES 8–9 , 15–16 View FIGURES 15–16 ).
Male tergopleurites II–IX+X and female tergopleurites II–VIII divided medianly; ventral sections of tergopleurites generally slender. Sternal plates rectangular, not approaching ventral sections of tergopleurites; accessory sternal plates absent (e.g. Figs 1–2 View FIGURES 1–2 , 8–9 View FIGURES 8–9 , 15–16 View FIGURES 15–16 ).
Male. Abdominal chaetotaxy sparse, differing slightly between species. Subgenital plate roughly triangular, lateral margins typically irregular (e.g. Figs 1 View FIGURES 1–2 , 8 View FIGURES 8–9 , 15 View FIGURES 15–16 ). Genitalia: basal apodeme rectangular, with rounded anterior end, often constricted at mid-length (e.g. Figs 4 View FIGURES 3–7 , 11 View FIGURES 10–14 , 18 View FIGURES 17–21 ). Proximal mesosome variable, typically quadratic or rectangular, narrow compared to distal mesosome; ventral sclerite present, variable between species; mesosomal lobes gener-ally wider than proximal mesosome, with distinct bulging nodi lateral to gonopore; these nodi are typically at least partially rugose; 2 ames sensilla and 2 pmes sensilla on each side (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ). Parameral heads variable (e.g. Figs 6 View FIGURES 3–7 , 13 View FIGURES 10–14 , 20 View FIGURES 17–21 ). Parameral blades tapering only distally, may be slightly elongated distal to mesosome; pst1 sensillus located proximal to pst2; pst2 microseta, near distal end of paramere (e.g. Figs 4, 6 View FIGURES 3–7 , 11, 13 View FIGURES 10–14 , 18, 20 View FIGURES 17–21 ).
Female. Abdominal chaetotaxy sparse, differing slightly among species. Subgenital plate broad in anterior half, narrowing in posterior half; lateral margins of posterior half often irregular; subgenital plate does not reach vulval margin, but expands distally into lateral submarginal bulges; few vms and vss on each side; vos follow lateral margins of subgenital plate, with at least 1 distal vos separated from other vos by a gap, and distal most vos typically situated on or near distal margin of subgenital plate, near vss (e.g. Figs 7 View FIGURES 3–7 , 14 View FIGURES 10–14 , 21 View FIGURES 17–21 ).
Host distribution. Species of Dicruridae , Oriolidae and Vangidae .
Geographical range. Afrotropical, Indo-Malayan and Australo-Papuan regions; presumably present outside these regions wherever drongos occur.
Etymology. The name Dicrurobates is formed by the host genus Dicrurus Vieillot, 1816 —from Greek “ dikros ” = “forked”, and “ oura ” = “tail”—combined with “ bates ”, Greek for “one who walks on something”.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Guimaraesiella ( Dicrurobates ) Gustafsson & Bush
| Gustafsson, Daniel R. & Bush, Sarah E. 2020 |
Brueelia
| Bush, S. E. & Weckstein, J. D. & Gustafsson, D. R. & Allen, J. & DiBlasi, E. & Shreve, S. C. & Boldt, R. & Skeen, H. R. & Johnson, K. P. 2016: 743 |
Brueelia Kéler, 1936: 257
| Keler, S. von 1936: 257 |